
Los fosfitos (Phi), derivados del ácido fosforoso, son compuestos químicos biocompatibles que han demostrado ser efectivos en el manejo integrado de cultivos (MIC). A diferencia de los fosfatos (Pi), los fosfitos no participan en rutas metabólicas esenciales debido a la ausencia del gen ptxD en las plantas, lo que evita su conversión en Pi. Sin embargo, poseen propiedades únicas que los convierten en potentes inductores de resistencia sistémica. Estimulan las defensas naturales de las plantas al activar genes de defensa, reforzar barreras estructurales y aumentar la producción de compuestos antimicrobianos. Además, interfieren en el metabolismo de los patógenos, dificultando su capacidad de inhibir las respuestas inmunológicas de las plantas y aumentando su visibilidad para un ataque más efectivo. Phi es altamente móvil en la planta, moviéndose tanto por xilema como floema, lo que facilita su aplicación eficiente. Su uso combinado con fungicidas ofrece un control integrado y sostenible
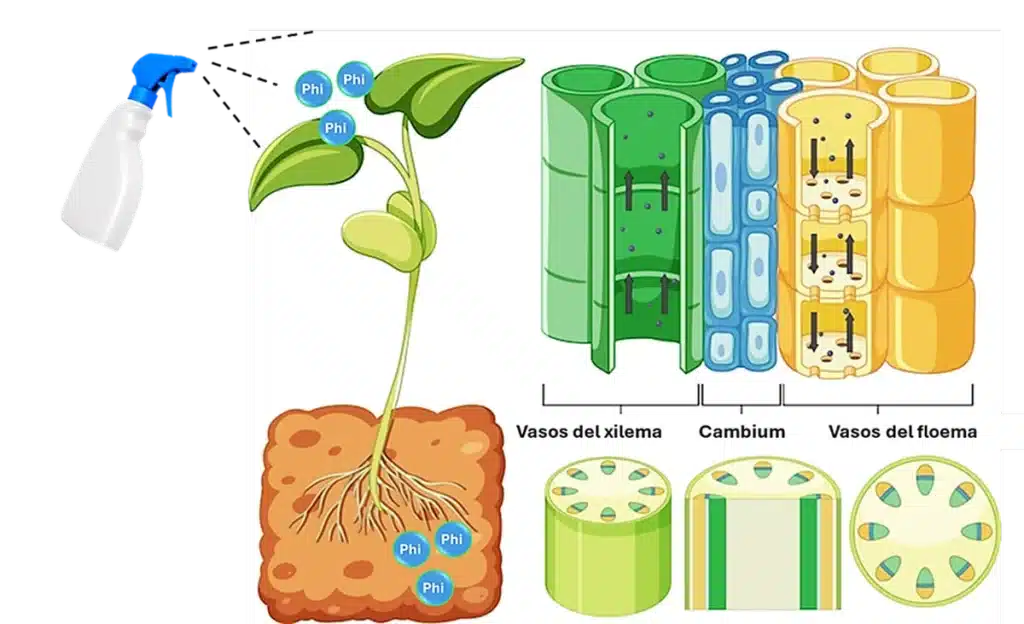
La búsqueda de soluciones sostenibles para las principales problemáticas de la agricultura como el manejo de enfermedades ha adquirido relevancia, ya que la comercialización agrícola actual demanda productos inocuos para los consumidores, provenientes de procesos con bajo impacto ambiental. La producción de más alimentos, sanos e inocuos, requiere necesariamente de la investigación e innovación como detonadores de nuevos mecanismos de los sistemas de producción, comercialización e industrialización de alimentos. Una estrategia innovadora, dentro del manejo integrado de cultivos (MIC), es el uso de compuestos químicos biocompatibles que potencian la resistencia de las plantas a enfermedades a través del mecanismo de resistencia inducida (RI) (Borza et al., 2017). Esta es una alternativa prometedora, que radica en la estimulación molecular de los mecanismos naturales de defensa de las plantas, siendo efectiva frente a una amplia gama de estreses (Daniel & Guest, 2006) .
Los fosfitos (Phi) son una forma reducida de los fosfatos (Pi), derivados del ácido fosforoso (H3PO3 – ), que regularmente se combinan con cationes no metálicos como potasio, sodio, calcio o amonio (Havlin & Schlegel, 2021). La diferencia química entre fosfato (H2PO4 – ; Pi) y fosfito (H2PO3 – ; Phi); es un átomo de oxígeno el cual es sustituido por otro de hidrógeno (Figura 1) (Lovatt & Mikkelsen, 2006). Tanto Pi como Phi muestran una geometría molecular tetraédrica, pero debido a la diferencia estructural, la distribución de la carga es distinta en cada molécula. Por lo tanto, tanto la forma de la molécula como la distribución de la carga parecen influir en la unión de Pi y Phi a sus moléculas interactuantes, y la mayoría de las enzimas involucradas en las reacciones de transferencia de fosforilo discrimina fácilmente entre Phi y Pi (Gómez-Merino & Trejo-Téllez, 2016).
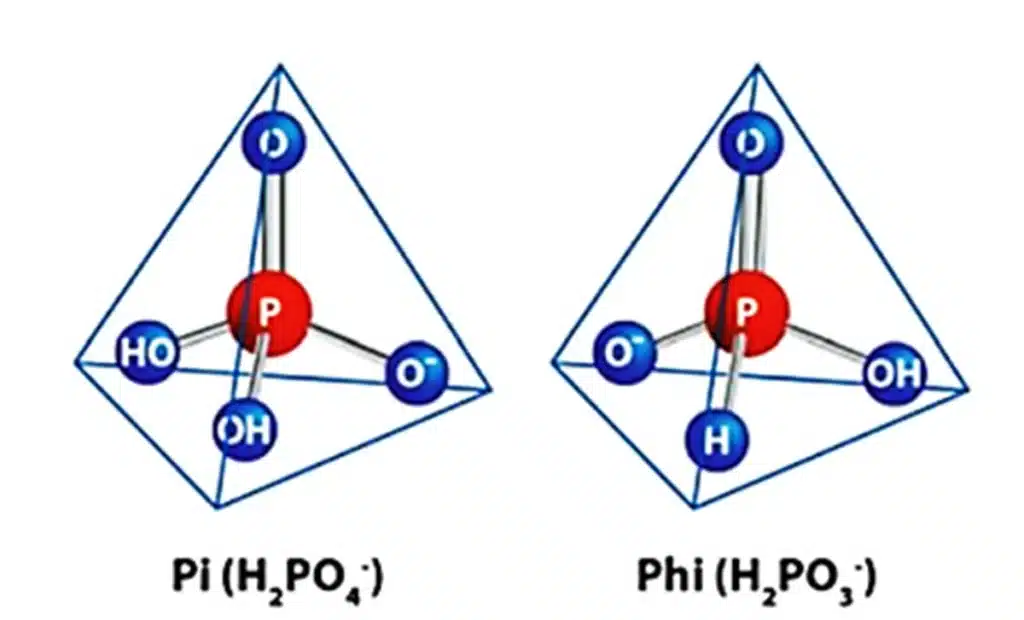
Los Phi no pueden ser convertidos en Pi dentro de la planta, debido a la ausencia del gen ptxD, el cual está presente en muchas bacterias de suelo (Wu et al., 2018); por lo tanto, no participan en las rutas bioquímicas, por ello se observan efectos negativos de estos sobre el metabolismo vegetal (Varadarajan et al., 2002). No obstante, los Phi hacen más eficaz el uso de fungicidas específicos e incrementan las defensas de las plantas ante la posibilidad de infección de algún patógeno (Bettiol, 2006; Lovatt y Mikkelsen, 2006).
Los fosfitos han sido ampliamente estudiados como una alternativa para el control de organismos fitoparásitos. Su eficacia se ha probado en contra de protozoarios, oomycetes de los géneros Peronospora, Plasmopara, Phytophthora y Pythium, hongos de los géneros Alternaria, Rhizoctonia y Macrophomina, bacterias y nematodos (Gómez-Merino & Trejo-Téllez, 2016).
Mecanismos de acción
La efectividad de los fosfitos en el control de fitopatógenos ocurre mediante vía directa e indirecta. En cuanto a la vía directa, el ion fosfito al entrar en contacto con los organismos fitopatógenos, afecta su crecimiento y reproducción, al inhibir la germinación de esporas al bloquear el desarrollo del micelio, compitiendo con el fosfato como regulador alostérico de varias enzimas causando una reducción en la síntesis de ATP, NAD. Como se describió anteriormente, Phi también compite por los sitios de unión de Pi de las enzimas fosforilantes. Por lo tanto, el efecto antifúngico de Phi en los oomicetos probablemente esté relacionado con la interferencia de Phi con el metabolismo de Pi (Havlin & Schlegel, 2021). A pesar de la información sobre el efecto de Phis en diferentes cultivos, su modo de acción no está claro y su molécula/s diana en la planta aún se desconoce (Feldman et al., 2020).
En relación con la vía indirecta, Feldman et al., 2020, expuso que el KPhi potencia la respuesta de defensa celular justo cuando es necesario. Es decir, ante un ataque de patógenos evitando un costo metabólico que podría afectar el rendimiento del cultivo. Se debe tener en cuenta que las defensas de las plantas al ataque de los patógenos son un complejo de reacciones químicas, las cuales se activan en el momento en que un patógeno penetra el tejido vegetal, en este momento la planta reconoce el invasor y activa alarmas internas para bloquear el ataque y frenar la infección (Madriz, 2002).Los Phi estimulan la inmunidad de la planta al aumentar la actividad de las defensas inducidas de la planta, como la producción de compuestos antimicrobianos, la activación de genes de defensa y el refuerzo de las paredes celulares.
En resumen, los fosfitos interfieren con el patógeno al reducir la capacidad del patógeno para inhibir las defensas de la planta. Esto podría deberse a que los fosfitos afectan la síntesis o la función de los efectores del patógeno. También aumentan la visibilidad del patógeno: Se ha observado que los fosfitos pueden alterar la actividad metabólica del patógeno, generando señales que lo hacen más detectable para la planta. Esto facilita una respuesta inmunológica más rápida y efectiva. En conjunto, los fosfitos actúan como inductores de resistencia sistémica en las plantas, ayudándolas a detectar y combatir infecciones de manera más eficiente. Sin embargo, su efectividad puede variar dependiendo del tipo de planta, del patógeno involucrado y de las condiciones ambientales. Por ello, suelen usarse como parte de un manejo integrado de enfermedades.
Absorción y transporte
Los Phi comparten los mismos transportadores de Pi tanto en la raíz como en las hojas. Como fungicida se puede mover de forma acropétala y basipétala. Su alta movilidad tanto vía floema como xilema le da una alta movilidad dentro de la planta, incluso se puede retraslocar (Gómez-Merino & Trejo-Téllez, 2015).
McDonald et al., 2001, propuso que Phi puede agravar la deficiencia de fosfato inorgánico al engañar a las células para que perciban que tienen suficiente Pi. Esto sugiere que la acumulación y posible toxicidad de Phi en las plantas podría deberse a una menor asimilación de Pi o a la incapacidad de metabolizar Phi o convertirlo en Pi dentro de las células. Además, Phi podría ser identificado por las plantas como si fuera Pi, lo que inhibiría las respuestas típicas de deficiencia de Pi necesarias para mantener el crecimiento y funcionamiento en suelos pobres en fósforo. Debido a su mayor solubilidad, el Phi se absorbe y transloca más rápidamente dentro de la planta que el Pi (Havlin & Schlegel, 2021).
Fosfito de potasio en acción
En plantas de pepino inoculadas con Pythium ultimum la aplicación de fosfito de potasio (KPhi) demostró que los tratamientos con 2 y 4 g L-1 podrían aliviar el daño de la enfermedad en gran medida, mientras que las plantas de control fueron severamente dañadas por el patógeno (Mofidnakhaei et al., 2016). En el cultivo de papa la aplicación preventiva de KPhi redujo la severidad del daño ocasionada por P. infestans. La planta aumentó su defensa estructural al aumentar los depósitos de calosa (Machinandiarena et al., 2012). Bajo condiciones in vitro, se evaluó el efecto de KPhi sobre P. cinnamomi, se observó un deterioro y una lisis de las células del patógeno debido a que KPhi puede afectar las enzimas relacionadas con la síntesis de celulosa. Esto último explica en parte por que KPhi tiene mayor eficacia contra oomycetes (King et al., 2010). En plantas de arroz (Oryza sativa). KPhi redujo la severidad de la pudrición del tallo del arroz causada por Nakataea oryzae (Catt.) J. Luo & N. Zhang. Las mezclas de Phi y fungicida dieron el mejor control de la pudrición del tallo e incrementaron el rendimiento. KPhi solo en dosis simples y dobles no protege contra la pudrición del tallo del arroz como fungicida. El KPhi puede integrarse en las estrategias de control de la pudrición del tallo del arroz (Martinez, 2016).
Perspectivas futuras
El uso de fosfitos en la agricultura presenta oportunidades para la innovación y la sostenibilidad. Las investigaciones futuras se enfocarán en optimizar las formulaciones para maximizar su efectividad y minimizar posibles impactos negativos, como la acumulación de Phi en suelos pobres en fósforo. Además, el estudio de sus interacciones con microbiomas vegetales puede ampliar su aplicación en estrategias de manejo integrado. Integrar fosfitos con tecnologías emergentes, como sensores para monitorear la salud de las plantas, podría mejorar la precisión en el manejo de cultivos. Finalmente, promover su uso en sistemas de producción orgánica y con certificaciones de bajo impacto ambiental fortalecerá su papel en la transición hacia una agricultura más sostenible y resiliente frente a los desafíos globales de seguridad alimentaria.
Existen alternativas a nivel comercial como, el fosfito de potasio EVOFERT PROTEK 32, el cual es un producto derivado del ácido fosforoso, diseñado para mejorar la resistencia de la planta frente a diversas enfermedades. EVOFERT PROTEK 32 es altamente sistémico, permitiendo su fácil movimiento tanto por xilema como en el floema llegando a todos los puntos de interés de la planta. Adicionalmente, EVOFERT PROTEK 32 inhibe el desarrollo del patógeno, afectando su crecimiento y reproducción, también advierte a las plantas del ataque de organismos patógenos para estimular la producción de fitoalexinas y los mecanismos de defensa natural.
Recomendaciones
La fitotoxicidad del fosfito depende del estado de nutrientes de P de las plantas y, por lo tanto, no se debe utilizar cuando las plantas tienen un estado de nutrientes de P subóptimo.

Descubre cómo potenciar la resistencia de tus cultivos con Protek32.
Gracias a su fórmula avanzada con fosfitos, activa las defensas naturales de las plantas y mejora su respuesta ante patógenos, asegurando cultivos más fuertes y productivos. Integra Protek32 en tu manejo agrícola y maximiza la protección con innovación y sostenibilidad. ¡Haz que tus plantas crezcan más sanas hoy mismo!
Referencias
- Bettiol, W. 2006. Productos alternativos para el manejo de enfermedades en cultivos comerciales. Fitosanidad. 10(2):85-98.
- Borza T, Peters RD, Wu Y, Schofield A, Rand J, Ganga Z, Al-Mughrabi KI, Coffin RH and Wang-Pruski G. 2017. Phosphite uptake and distribution in potato tubers following foliar and postharvest applications of phosphite-based fungicides for late blight control. Annals of Applied Biology 1:127-139. https://doi.org/10.1111/aab.12322
- Daniel R and Guest D. 2006. Defence responses induced by potassium phosphonate in Phytophthora palmivora-challenged Arabidopsis thaliana. Physiologycal and Molecular Plant Pathology 67:194-201.
- Feldman, M. L., Guzzo, M. C., Machinandiarena, M. F., Rey-Burusco, M. F., Beligni, M. V., Di Rienzo, J., Castellote, M. A., Daleo, G. R., & Andreu, A. B. (2020). New insights into the molecular basis of induced resistance triggered by potassium phosphite in potato. Physiological and Molecular Plant Pathology, 109, 101452. https://doi.org/10.1016/j.pmpp.2019.101452
- Gómez-Merino, F. C., & Trejo-Téllez, L. I. (2015). Biostimulant activity of phosphite in horticulture. Scientia Horticulturae, 196, 82–90. https://doi.org/10.1016/j.scienta.2015.09.035
- Gómez-Merino, F. C., & Trejo-Téllez, L. I. (2016). Conventional and novel uses of phosphite in horticulture: potentialities and challenges. Italus Hortus, 23(2), 1–13.
- Havlin, J. L., & Schlegel, A. J. (2021). Review of Phosphite as a Plant Nutrient and Fungicide. Soil Systems, 5(3), 52. https://doi.org/10.3390/soilsystems5030052
- King, M., Reeve, W., Van der Hoek, M. B., Williams, N., McComb, J., O’Brien, P. A., & Hardy, G. E. S. J. (2010). Defining the phosphite-regulated transcriptome of the plant pathogen Phytophthora cinnamomi. Molecular Genetics and Genomics, 284, 425-435.
- Lovatt, C. and Mikkelsen, R. 2006. Phosphite fertilizers: what are they? can you use them? what can they do? Better Crops. 90(4):11-13.
- Machinandiarena, M. F., Lobato, M. C., Feldman, M. L., Daleo, G. R., & Andreu, A. B. (2012). Potassium phosphite primes defense responses in potato against Phytophthora infestans. Journal of Plant Physiology, 169(14), 1417–1424. https://doi.org/10.1016/j.jplph.2012.05.005
- McDonald, A. E., Grant, B. R., & Plaxton, W. C. (2001). Phosphite (phosphorous acid): its relevance in the environment and agriculture and influence on plant phosphate starvation response. Journal of plant nutrition, 24(10), 1505-1519.
- Mofidnakhaei, M., Abdossi, V., Dehestani, A., Pirdashti, H., & Babaeizad, V. (2016). Potassium phosphite affects growth, antioxidant enzymes activity and alleviates disease damage in cucumber plants inoculated with Pythium ultimum. Archives of Phytopathology and Plant Protection, 49(9–10), 207–221. https://doi.org/10.1080/03235408.2016.1180924.
- Varadarajan, D. K; Karthikeyan, A. S.; Matilda, P. D. and Raghothama, K. G. 2002. Phosphite, an analog of phosphate, suppresses the coordinated expression of genes under phosphate starvation. Plant Physiol. 129(3):1232-1240.
- Wu, L.; Gao, X.; Xia, F.; Joshi, J.; Borza, T. and Wang-Pruski, G. 2018. Biostimulant and fungicidal effects of phosphite assessed by GC-TOF-MS analysis of potato leaf metabolome. Physiol. Mol. Plant Pathol. 106(1):49-56. https://doi.org/10.1016/j.pmpp. 2018.12.001.