 Saltar al contenido
Saltar al contenido
Transporte de azúcares en la planta y relación fuente vertedero
Definiciones:
- Fuente: órganos que producen más carbohidratos de los que consumen (ej. hojas maduras).
- Vertedero: órganos que consumen más carbohidratos de los que producen (ej. raíces, frutos, hojas jóvenes).
El transporte de azúcares en las plantas es un proceso fundamental que asegura la distribución de energía y nutrientes desde los órganos productores, conocidos como fuentes, hacia los órganos consumidores o de almacenamiento, llamados sumideros o vertederos.
Los azúcares se producen principalmente en las hojas mediante la fotosíntesis, siendo el producto principal la sacarosa, que es la forma principal de transporte de azúcares en muchas plantas. Este flujo desde zonas de alta concentración en las hojas hacia zonas de baja concentración en los sumideros es crucial para el crecimiento, la reproducción y el almacenamiento de reservas en diferentes partes de la planta, como raíces, frutos y flores.
La relación fuente-vertedero es dinámica y varía a lo largo del ciclo de vida de la planta, siendo influenciada por factores como el desarrollo de la planta, las condiciones ambientales y la regulación hormonal. Por ejemplo, en etapas tempranas de crecimiento, las hojas jóvenes actúan como sumideros hasta que maduran y comienzan a producir azúcares. Durante la fructificación, los frutos se convierten en los principales sumideros, atrayendo gran parte de los azúcares producidos. Las hormonas vegetales, como auxinas y citoquininas, también juegan un papel en la modulación de esta relación, afectando la capacidad de los órganos sumideros para atraer y utilizar los azúcares, lo que tiene implicaciones directas en la productividad y calidad de los cultivos.
Dinámica de la relación
- Fuerza del vertedero: Capacidad de un órgano para atraer fotoasimilados.
- Proximidad: Los vertederos más cercanos a la fuente suelen tener prioridad.
- Etapa de desarrollo: Las necesidades cambian según la etapa de crecimiento de la planta.
Transporte de azúcares y desarrollo de la planta
Los transportadores de carbohidratos se encuentran en casi todas las membranas de la célula y median el transporte de azúcares a través de la membrana plasmática. Las proteínas transportadoras de azúcar juegan un papel crucial en la regulación y están asociadas con diversos procesos de desarrollo en las plantas, por ejemplo, para controlar el desarrollo de órganos receptores como las raíces o los frutos, por lo cual se cuenta con casi 90 transportadores de azúcares caracterizados. Estos procesos, como la transición de la fase vegetativa a la reproductiva, la ramificación de brotes, la ramificación y elongación de raíces, el desarrollo de frutos y, finalmente, la senescencia, son influenciados por distintas proteínas que detectan y transportan azúcares a nivel celular. (Guo et al., 2023).
La planta usa transportadores de azúcares específicos para los procesos antes mencionados. Por ejemplo: el transportador de azúcar SWEET17 y TST1/2 (ubicados en la vacuola) transportan azúcares para la extensión y ramificación de raíces. Los transportadores de sacarosa Psut Y MEX1 ubicados en el cloroplasto, exportan azúcares para la ramificación de brotes aéreos. Avances recientes han revelado la función clave de las proteínas SWEET en la descarga inicial de sacarosa para el desarrollo del fruto, como también se informó para el desarrollo de semillas y polen (Guo et al., 2023). Los azúcares, especialmente la glucosa y la fructosa, contribuyen al sabor y la calidad de los frutos del tomate. Estos compuestos se translocan desde las hojas hasta los frutos y luego se descargan en los frutos mediante varios transportadores de azúcar en la membrana plasmática. SlSWEET7a y SlSWEET14, localizados en la membrana plasmática, regulan el transporte y almacenamiento de azúcar en frutos de tomate (Zhang et al., 2021).
En la reproducción sexual y, especialmente, durante el llenado de semillas, la senescencia representa un proceso controlado asociado con una importante transferencia de carbono y aminoácidos a las semillas en desarrollo (Guo et al., 2023). Durante el desarrollo del fruto, en el apoplasma vascular, hasta un 70% de Sacarosa (Sac) puede ser hidrolizada por la invertasa de la pared celular (CWIV por sus siglas en inglés) en glucosa y fructosa, que deben importarse rápidamente al parénquima de almacenamiento adyacente en el fruto para mantener el gradiente de sacarosa entre el elemento tamiz/Célula acompañante (ET/CA) para un flujo continuo de Sac a los vertederos (Ruan, 2014).
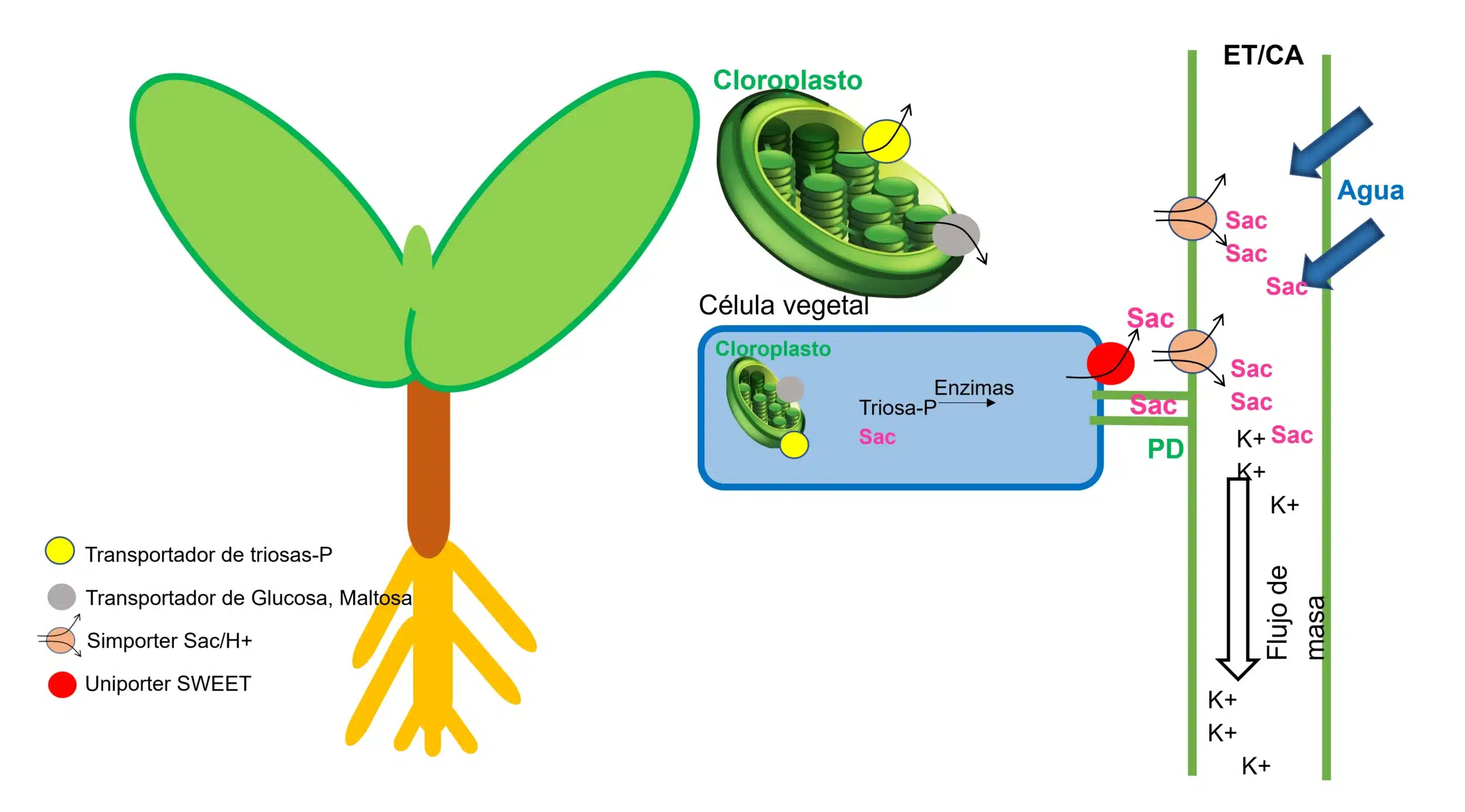
Imagen 1: Síntesis de sacarosa (Sac) y carga de floema en hojas fuente. Las hojas fotosintéticas (la fuente) usan energía solar para fijar CO2 y producir triosa-fosfato (triosa-P) en el cloroplasto. La triosa-P se exporta al citoplasma como componentes básicos para otros metabolismos: almidón, sacarosa, respiración celular, etc. La Sac se carga en el floema ya sea apoplásmicamente (a través de la pared mediado por transportadores) o simplásmicamente (a través de plasmodesmos (PD) que conectan el complejo ET/CA). La acumulación de Sac atrae agua osmóticamente, creando una alta presión de turgencia, que impulsa el flujo de masa de asimilados hacia los sumideros. K+: potasio, ET/CA: elemento tamiz/célula acompañante. Elaboración propia a partir de (Ruan, 2014).
Los gradientes de azúcar en la planta funcionan como formas de señalización para la expresión de genes. De hecho, la sacarosa puede funcionar como un regulador positivo o negativo en la expresión de genes. Bajo condiciones de escasez, las plantas activan mecanismos de respuesta que incluyen la movilización de reservas de carbohidratos y la regulación de la expresión génica para mitigar el impacto del estrés. Estos mecanismos son esenciales para la supervivencia en ambientes adversos y ofrecen una perspectiva para mejorar la tolerancia al estrés en cultivos mediante la biotecnología.
Papel de la vacuola en el transporte de azúcares: En la célula vegetal, la vacuola es el principal sitio de almacenamiento de azúcares (Hedrich et al., 2015). La membrana vacuolar o tonoplasto cuenta con un transportador de monosacáridos TST por sus siglas en inglés (glucosa/fructosa), y es uno de los elementos en la detección de azúcar celular. En Arabidopsis regula el rendimiento de la semilla.
En cuanto a la carga del floema, en un estudio hecho en maíz (Zea mays) (Babst et al., 2022) investigaron si la carga de azúcar en el floema es esencial para el flujo de savia en plantas de maíz. Tradicionalmente, se ha creído que la carga de azúcares, como la sacarosa, es necesaria para generar la presión osmótica que impulsa este flujo. Sin embargo, el estudio muestra que en un mutante de maíz con una función reducida del transportador de sacarosa (sut1), aunque la exportación de carbono se reduce drásticamente, la presión en el floema y el flujo de savia se mantienen en niveles significativos. Esto sugiere que otros factores, como el aumento de potasio (K+), pueden compensar la disminución de azúcar, permitiendo el flujo de savia. Los resultados indican que el flujo de savia y el transporte de otros nutrientes podrían regularse independientemente de la carga de azúcar, afectando la homeostasis de nutrientes y la distribución de señales en las plantas bajo diferentes condiciones ambientales.
Papel de algunos elementos en el transporte de azúcares
El potasio (K) se utiliza como osmolito para facilitar la carga de sacarosa en el floema, promover la síntesis de productos fotosintéticos y mantener el contenido de agua de las células (Shen et al., 2019). El K tiene la función de mantener el gradiente electroquímico de la membrana, la presión del floema y servir como fuente de energía móvil necesaria para la carga y transporte de asimilados en el floema. En un estudio hecho por Wu et al., (2023) en plantas de tomate fertilizadas con cloruro de potasio KCl, disminuyó el ácido málico (MA) y el ácido cítrico.(CA), pero aumentaron las concentraciones de fructosa (Fru), glucosa (Glu) y sacarosa (Suc) en las manzanas en la etapa de madurez. El fertilizante sulfato de potasio K2SO4 aumentó las concentraciones de Glu y Suc, pero no tuvo efectos sustanciales sobre las concentraciones de Fru y ácido titulable (TA) en peras asiáticas. El fertilizante K2SO4 aumentó las concentraciones de Suc, Glu, Fru y CA en frutos de naranja navel. El fertilizante KCl es mejor que el K2SO4 en la formación del rendimiento y la calidad de la fruta en la etapa de madurez, mientras que el fertilizante K2SO4 mejora el crecimiento de las plantas y prolonga la vida útil de las manzanas (Wu et al., 2023).
Se ha investigado cómo la deficiencia de fosfato, un nutriente clave, afecta la movilización de azúcares, revelando interacciones complejas entre la disponibilidad de nutrientes y la regulación del metabolismo de los carbohidratos, lo cual es fundamental para la eficiencia en el uso de nutrientes en las plantas.
El efecto del boro (B) sobre los tejidos jóvenes en crecimiento demuestra su papel crítico principalmente en la división y elongación celular. También se reporta que el B influye en la disponibilidad y absorción de otros nutrientes vegetales del suelo, como el aumento aparente en la absorción y translocación de P, N, K, Zn, Fe y Cu en hojas, brotes y semillas después de la aplicación de B en algodón. Inicialmente, se ha argumentado que la difusión pasiva de ácido bórico sin carga a través de la bicapa lipídica es el único mecanismo de transporte de B desde las raíces de las plantas vasculares hasta las partes aéreas de la planta, como el tallo y las hojas (Shireen et al., 2018). El boro juega un papel importante en la mejora del tamaño y número de células en las plantas (Bredun et al., 2021).
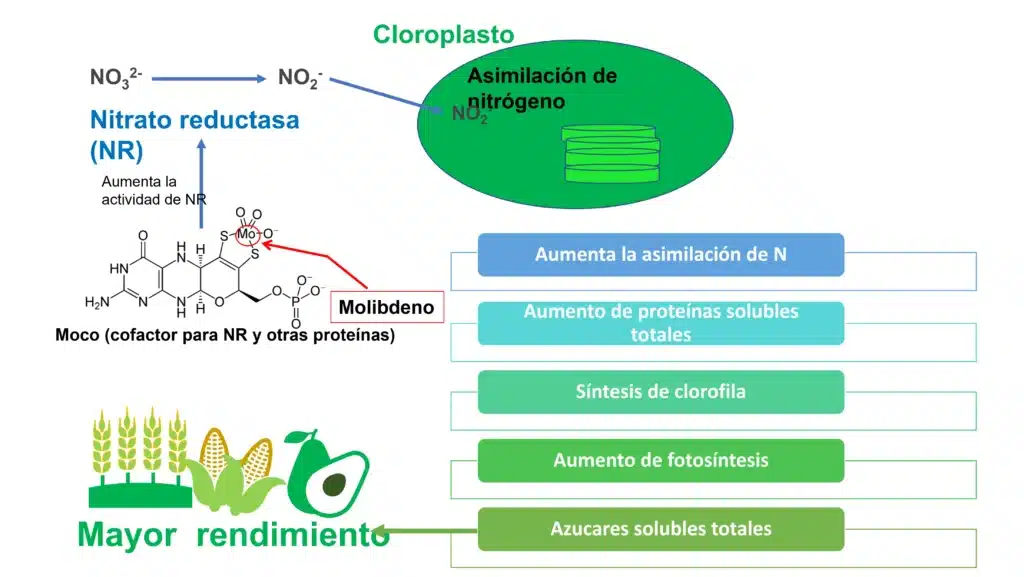
La forma en que la planta absorbe el Mo es molibdato (MoO4-2) y su absorción se ve favorecida con un pH ˃ 5.5 (Alamri et al., 2022; Vigani et al., 2017). Para adquirir actividad biológica en la célula, el Mo tiene que ser incorporado a una molécula denominada en los diferentes artículos consultados como un andamio de pterina para formar el cofactor de molibdeno (Moco) (Mendel, 2022). Como se puede ver en la figura 1, y la tabla 2 esta nueva molécula que contiene Mo es cofactor de enzimas tan importantes como la nitrato reductasa NR, nitrito reductasa NiR indispensables en el metabolismo del N dentro de la célula vegetal. Por último, el Mo parece estar implicado en la formación del ABA al ser parte estructural de la enzima que genera el ácido abscísico (Azcón & Talón, 2013).
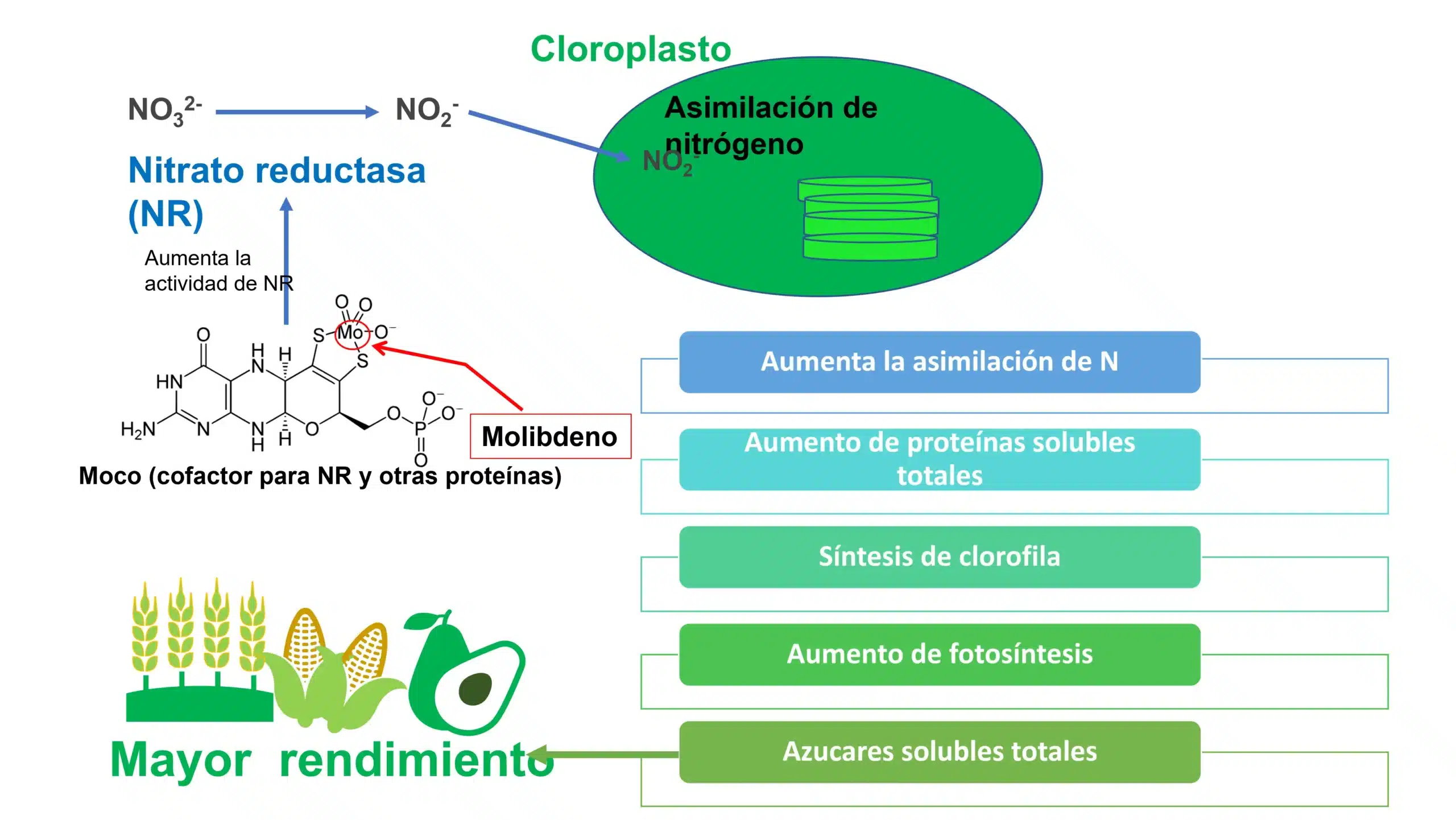
Figura 2: Función indirecta del molibdeno en la movilización de azúcares. La nitrato reductasa (NR) es una enzima que cataliza la reducción de nitrato (NO32-) a nitrito (NO2-). Es usuaria del cofactor de molibdeno MOCO que contiene molibdeno. Mo también actúa directamente sobre la fase fotoquímica de la fotosíntesis a través de la biosíntesis de clorofila y la estabilidad del aparato fotosintético. El aumento de la actividad fotosintética activa la NR y promueve el inicio de la traducción de proteínas. Por lo tanto, hay un aumento en el contenido de proteína soluble total en las hojas, lo que puede conducir a un aumento en el contenido y la actividad de la enzima rubisco. Esta enzima es responsable de la fijación de carbono durante la producción de carbohidratos, lo que conduce a un aumento en el contenido de azúcares solubles totales en las plantas, y finalmente mayor rendimiento. Elaboración propia a partir de (Bittner, 2014; Oliveira et al., 2022).
La actividad del transportador de azúcar es fundamental para el desarrollo adecuado de los órganos, lo que convierte a las proteínas transportadoras de azúcar en objetivos interesantes para la modificación y el estudio de la influencia de la distribución de carbohidratos en el desarrollo de las plantas. El transporte de azúcares en las plantas no solo es vital para el crecimiento y desarrollo, sino que también está intrínsecamente ligado a la regulación hormonal y a la influencia de nutrientes clave como el potasio, el boro y el molibdeno. La capacidad de los vertederos para atraer y utilizar los azúcares está modulada por transportadores específicos que garantizan la adecuada distribución de carbohidratos en la planta. Además, la dinámica de la relación fuente-vertedero y la interacción con elementos esenciales tiene un impacto directo en la productividad y calidad de los cultivos, subrayando la importancia de una gestión adecuada de los nutrientes y el monitoreo del desarrollo vegetal para optimizar el rendimiento agrícola.

Máximo rendimiento con Sugar Flow: Boro y Molibdeno en Acción
Sugar Flow es un producto diseñado para incrementar la movilización de azúcares en el interior de la planta. La tecnología AdGen de esta formulación promueve el transporte de azúcares de la fuente (hoja) al sumidero u órgano de interés (fruto, tubérculo, grano). Además de azúcares y nutrientes, es precursor en la síntesis y transporte de fitohormonas.
En leguminosas y cereales se aplica durante el desarrollo de la semilla, en etapa de grano lechoso. Sugar Flow aumenta la fuerza de los vertederos, promueve la acumulación de masa seca en los órganos de interés, mejora la calidad de los frutos: aumenta la concentración de azúcares, SST, ácidos titulables. Además, Sugar Flow puede proporcionar B y Mo si se encuentran deficiencias en la planta. Finalmente, el contenido de B y Mo pueden conferir a la planta tolerancia a periodos de estrés abiótico (Pandey, 2018).
Referencias
Alamri, S., Siddiqui, M. H., Mukherjee, S., Kumar, R., Kalaji, H. M., Irfan, M., Minkina, T., & Rajput, V. D. (2022). Molybdenum-induced endogenous nitric oxide (NO) signaling coordinately enhances resilience through chlorophyll metabolism, osmolyte accumulation and antioxidant system in arsenate stressed-wheat (Triticum aestivum L.) seedlings. Environmental Pollution, 292. https://doi.org/10.1016/j.envpol.2021.118268
Azcón, J., & Talón, M. (2013). Fundamentos de Fisiologia Vegetal: Vol. I (Segunda edición). McGraw-Hill Interamericana. https://www.facebook.com/recursos.para.agronomos.chapingo
Babst, B. A., Braun, D. M., Karve, A. A., Frank Baker, R., Tran, T. M., Kenny, D. J., Rohlhill, J., Knoblauch, J., Knoblauch, M., Lohaus, G., Tappero, R., Scherzer, S., Hedrich, R., & Jensen, K. H. (2022). Sugar loading is not required for phloem sap flow in maize plants. Nature Plants, 8(2), 171–180. https://doi.org/10.1038/s41477-022-01098-x
Bittner, F. (2014). Molybdenum metabolism in plants and crosstalk to iron. In Frontiers in Plant Science (Vol. 5, Issue FEB). Frontiers Research Foundation. https://doi.org/10.3389/fpls.2014.00028
Bredun, M. A., Gomes, T. M., Assumpção, T. I., Brighenti, A. F., Chaves, E. S., Panceri, C. P., & Burin, V. M. (2021). Statement of Boron application impact on yield, composition and structural properties in Merlot grapes. Scientia Horticulturae, 288. https://doi.org/10.1016/j.scienta.2021.110364
Guo, W.-J., Pommerrenig, B., Neuhaus, H. E., & Keller, I. (2023). Interaction between sugar transport and plant development. Journal of Plant Physiology, 288, 154073. https://doi.org/10.1016/j.jplph.2023.154073
Hedrich, R., Sauer, N., & Neuhaus, H. E. (2015). Sugar transport across the plant vacuolar membrane: nature and regulation of carrier proteins. Current Opinion in Plant Biology, 25, 63–70. https://doi.org/10.1016/j.pbi.2015.04.008
Mendel, R. R. (2022). The History of the Molybdenum Cofactor—A Personal View. In Molecules (Vol. 27, Issue 15). MDPI. https://doi.org/10.3390/molecules27154934
Oliveira, S. L., Crusciol, C. A. C., Rodrigues, V. A., Galeriani, T. M., Portugal, J. R., Bossolani, J. W., Moretti, L. G., Calonego, J. C., & Cantarella, H. (2022). Molybdenum Foliar Fertilization Improves Photosynthetic Metabolism and Grain Yields of Field-Grown Soybean and Maize. Frontiers in Plant Science, 13. https://doi.org/10.3389/fpls.2022.887682
Ruan, Y. L. (2014). Sucrose metabolism: Gateway to diverse carbon use and sugar signaling. In Annual Review of Plant Biology (Vol. 65, pp. 33–67). Annual Reviews Inc. https://doi.org/10.1146/annurev-arplant-050213-040251
Shen, C., Shi, X., Xie, C., Li, Y., Yang, H., Mei, X., Xu, Y., & Dong, C. (2019). The change in microstructure of petioles and peduncles and transporter gene expression by potassium influences the distribution of nutrients and sugars in pear leaves and fruit. Journal of Plant Physiology, 232, 320–333. https://doi.org/10.1016/j.jplph.2018.11.025
Shireen, F., Nawaz, M. A., Chen, C., Zhang, Q., Zheng, Z., Sohail, H., Sun, J., Cao, H., Huang, Y., & Bie, Z. (2018). Boron: Functions and approaches to enhance its availability in plants for sustainable agriculture. In International Journal of Molecular Sciences (Vol. 19, Issue 7). MDPI AG. https://doi.org/10.3390/ijms19071856
Vigani, G., di Silvestre, D., Agresta, A. M., Donnini, S., Mauri, P., Gehl, C., Bittner, F., & Murgia, I. (2017). Molybdenum and iron mutually impact their homeostasis in cucumber (Cucumis sativus) plants. New Phytologist, 213(3), 1222–1241. https://doi.org/10.1111/nph.14214
Wu, K., Hu, C., Wang, J., Guo, J., Sun, X., Tan, Q., Zhao, X., & Wu, S. (2023). Comparative effects of different potassium sources on soluble sugars and organic acids in tomato. Scientia Horticulturae, 308, 111601. https://doi.org/10.1016/j.scienta.2022.111601Zhang, X., Feng, C., Wang, M., Li, T., Liu, X., & Jiang, J. (2021). Plasma membrane-localized SlSWEET7a and SlSWEET14 regulate sugar transport and storage in tomato fruits. Horticulture Research, 8(1), 186. https://doi.org/10.1038/s41438-021-00624-w