
El aguacate (Persea americana Mill) es originario de América. Hay evidencia arqueológica de su uso y selección en México desde hace 10.000 años. El aguacate, domesticado por los Aztecas, se dispersó desde México hasta el Perú en el período precolombino. Después del descubrimiento de América, y con la conquista fue llevado a España en 1600 y, posteriormente, comenzó su distribución a nivel mundial: Jamaica, Cuba, Ghana, etc. En Colombia, el aguacate puede crecer desde 0 hasta 2.500 m s. n. m., en zonas de cordillera, lo que pone de manifiesto que no se conocen del todo las mejores condiciones agroclimáticas para su crecimiento [1].
A nivel mundial los países con mayor área sembrada y producción son: México, Colombia, Perú, Indonesia, Chile, Sur África y EE. UU. Los países con mayor rendimiento en los últimos años son: El Salvador, Panamá y Samoa (figura 1). En Colombia, el cultivo ha crecido en área, producción y rendimiento en los últimos años, en el año 2020 se sembraron 79.832 hectáreas con una producción 889.866 toneladas y un rendimiento de 11,15 ton/ha. Actualmente los departamentos más representativos son Antioquia, Tolima, Caldas, Valle del Cauca y Santander [2].
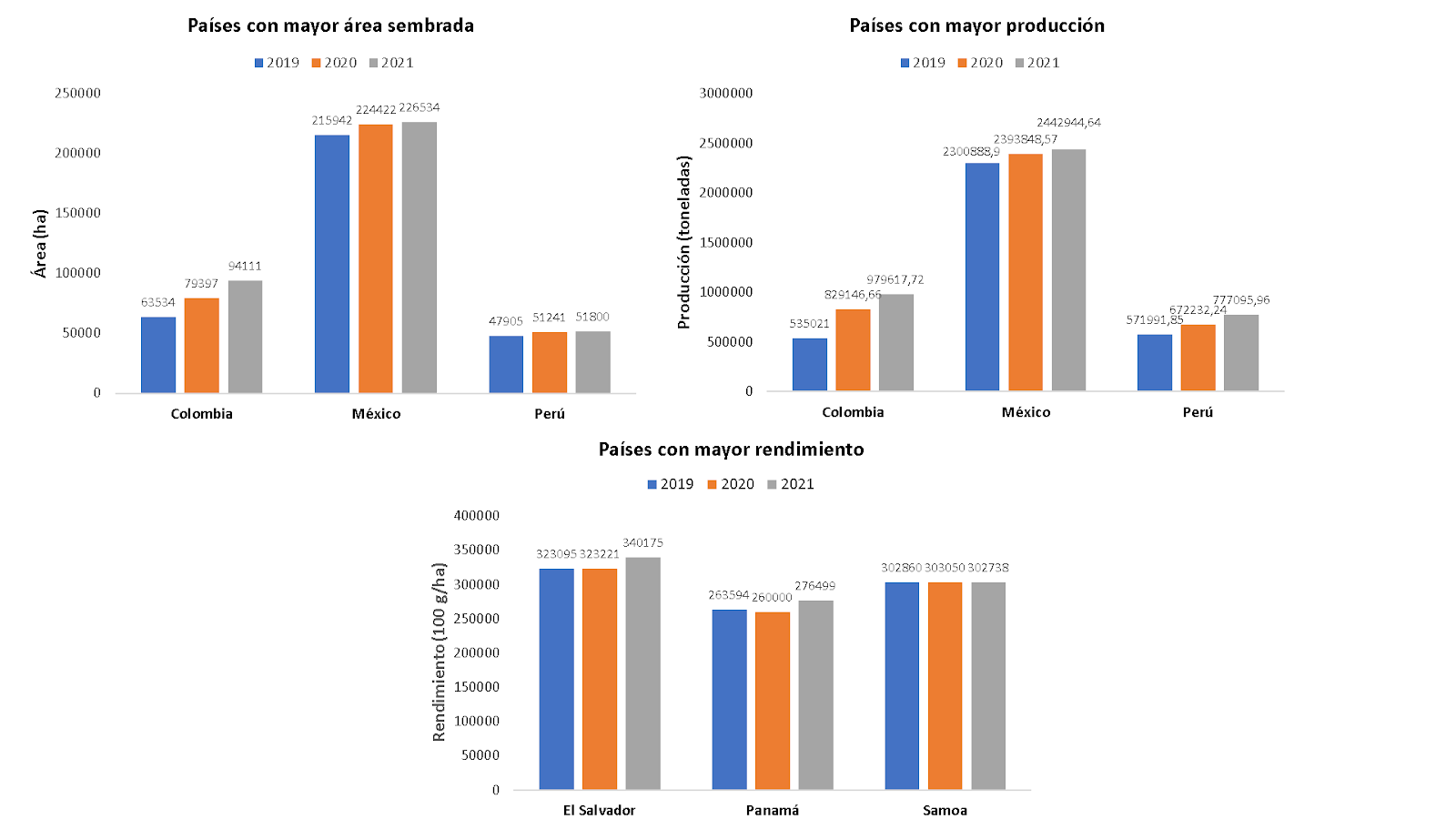
Figura 1: países con mayor área sembrada producción y rendimiento de aguacate en el mundo entre el año 2019 y 2021. Elaboración propia a partir de datos obtenidos de FAO, (2023).
Un árbol de aguacate puede llegar a producir entre 1 y 2 millones de flores, tan solo el 1 al 0.01% de las flores llegan a convertirse en frutos, lo que equivale a 100 a 200 frutos/árbol. El aguacate puede presentar alternancia en su producción, es decir periodos de muy buena producción seguido de un periodo de baja producción [1]. El aguacate tiene dos tipos de brotes, indeterminados, que mantienen el desarrollo vegetativo a partir de una yema apical, y determinados, que no tienen crecimiento vegetativo. Los brotes indeterminados dan menos frutos que los determinados y aceleran significativamente la abscisión fisiológica de los frutos.
D’Asaro y colaboradores [4], notaron que, en la etapa de antesis y cuajado, las concentraciones de giberelinas y citoquininas (principalmente GA 1 y t Z, respectivamente) fueron significativamente mayores en las flores de inflorescencias determinadas que en las indeterminadas, lo que indica su mayor capacidad de cuajado. Se ha demostrado que una mayor concentración de hormonas como citoquininas y giberelinas están relacionadas con una mayor retención de flores y de frutos.

Lo anterior es importante porque el rendimiento promedio mundial es de aproximadamente 10 ton/ha y en Colombia 14 ton/ha independiente de la alternancia. Esta baja producción no se debe a una floración deficiente, como ya se mencionó un árbol de aguacate produce cientos de miles de flores en una etapa determinada, por lo que la baja producción está relacionada es con la alta abscisión o caída de flores y frutos [1].
Además, Garner y colaboradores [5] proporcionan una fuerte evidencia de que la mayoría de las flores y frutos del aguacate ‘Hass’ se caen debido a la falta de germinación del polen y la subsiguiente fertilización. Sus resultados sugieren que la acumulación de ABA está relacionada con el aborto de óvulos (semillas) y la reducción del crecimiento de la fruta en la abscisión de la fruta.
Por otro lado, Moor y colaboradores [6] establecieron que en aguacate el parámetro limitante para el crecimiento de la variante de fruta pequeña parece ser el número de células, y se demostró que se produce una reducción en el número de células que coincide con el aumento de la concentración de ABA en el mesocarpio y la reducción de la actividad de 3-hidroxi-3-metilglutaril coenzima A reductasa (HMGR). El retraso del crecimiento de la fruta inducido por ABA y la inhibición de la actividad de HMGR son anulados por el tratamiento conjunto con Isopentiladenina (iPA) una citoquinina (CK), se sugirió una relación entre la actividad de HMGR y la concentración endógena de ABA y CK en el control metabólico del crecimiento de la fruta de aguacate Hass.
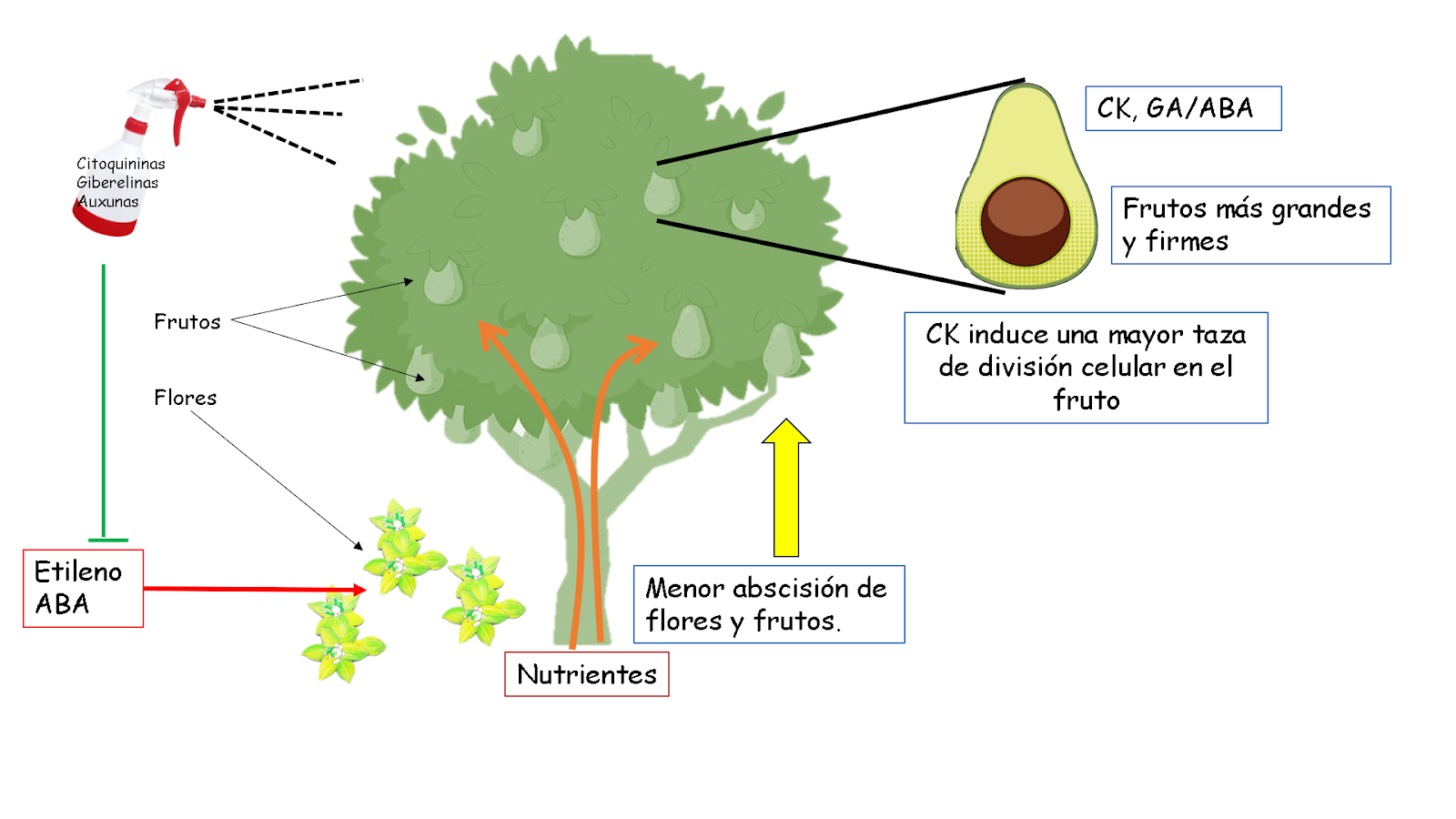
Figura 2: efecto de la aplicación de fitohormonas sobre la retención de flores y frutos, cuajado, tamaño y número de frutos. La aplicación foliar de auxinas, citoquininas y giberelinas mejora la absorción de nutrientes, disminuye el aborto de flores y frutos, promueve la división celular, la fuerza de llenado del fruto por lo que cada planta produce una mayor cantidad de frutos de un mejor calibre. Las auxinas aumentan la fuerza de vertedero de la fruta y regula el desarrollo del endospermo. Una alta concentración de etileno y ABA durante el periodo de floración y cuajado de frutos está relacionado con una mayor abscisión de estos. La prevención del pico de etileno y la abscisión asociada de los frutos podría, por lo tanto, estar bajo el control de otras sustancias de crecimiento vegetal como las citoquininas, que muestran un fuerte pico durante este período. Un cambio en el balance de las hormonas de crecimiento y las inhibidoras relacionadas con el estrés podría reducir la abscisión de flores y frutos.
Se ha demostrado que la aplicación de hormonas vegetales en etapas estratégicas del cultivo puede mejorar la floración, el amarre de frutos y por tanto, el rendimiento. En un artículo anterior del foro: ¿Qué son las fitohormonas y cómo funcionan?, se menciona la función que cumplen estos biorreguladores en la planta. Estas hormonas regulan y controlan una amplia variedad de procesos de crecimiento y desarrollo. En el caso del aguacate, las fitohormonas son fundamentales para varios aspectos del ciclo de vida y la producción de la fruta. En el aguacate, las citoquininas pueden estimular la formación de brotes laterales, promover la floración y mejorar el cuajado de frutos. A continuación, se destacan algunas funciones adicionales de las citoquininas y su relevancia:
Las citoquininas son una clase de hormonas vegetales que trabajan junto con las auxinas para regular el crecimiento y desarrollo de las plantas. Las citoquininas son producidas en las raíces y en los tejidos de los brotes jóvenes, y se transportan a través del xilema a otras partes de la planta.
- Estimulan la división celular: Las citoquininas promueven la división celular en las plantas, lo que les permite crecer y desarrollarse. Estas hormonas interactúan con las auxinas para controlar el crecimiento de la planta, regulando la relación entre la tasa de división celular y la tasa de elongación celular.
- Retrasan el envejecimiento: Las citoquininas también tienen un efecto antienvejecimiento en las plantas, ya que pueden retrasar la senescencia o envejecimiento de las hojas y otros órganos vegetales. Al inhibir la degradación de proteínas y la formación de radicales libres, las citoquininas pueden mantener las células y tejidos vegetales jóvenes y saludables durante más tiempo.
- Promueven la ramificación y la formación de brotes laterales: Las citoquininas pueden estimular la formación de brotes y ramas laterales en las plantas, lo que aumenta la capacidad de la planta para capturar la luz solar y producir más flores y frutos.
- Regulan la transpiración: Las citoquininas también pueden regular la tasa de transpiración de las plantas, ayudando a conservar el agua en condiciones de sequía.
En general, las citoquininas son hormonas importantes en las plantas que regulan el crecimiento y desarrollo de la planta, promoviendo la división celular, retrasando el envejecimiento, promoviendo la ramificación y regulando la transpiración. Las citoquininas trabajan en conjunto con otras hormonas vegetales, como las auxinas, para mantener un equilibrio adecuado de crecimiento y desarrollo en la planta.
| Tipo de citoquinina | Concentración | Tipo y forma de aplicación | Respuesta fisiológica | Autor |
| Isopentilade-nina (iPA) | 1 mg/mL | Inyectado en pedicelo del fruto. | Revirtió el efecto de ABA y mevastatina en la detención del crecimiento del fruto. | [7] |
| iPA | 1 mg/mL | Inyectado en pedicelo del fruto. | La aplicación de ABA causó senescencia en la cubierta de la semilla y retraso en el crecimiento de la fruta, pero estos efectos estuvieron ausentes en la fruta tratada con cantidades iguales de ABA más iPA. | [6] |
| Benciladenina, kinetina, Ipa, zeatina, encymidol (similar a ck) | 500 µM. 0.6 mL volumen | La fruta se partido por mitad se extrajo el mesocarpio y se le aplicaron las CK. | Las citoquininas, bencil adenina, kinetina, isopentenil adenina y zeatina y el análogo de citoquinina, ancymidol, inhibieron la biosíntesis de ácido abscísico a partir de ácido mevalónico en el tejido del mesocarpio de aguacate. | [8] |
| NAA + CPPU | NAA: 15 y 30 ppmCPPU 5 y 10 ppm | Aspersión foliar en plena floración y comienzo de fructificación (15 días después) | Los árboles de aguacate tratados con 30 ppm de NAA + 5 ppm de CPPU dieron como resultado el mayor cuajado de frutos/panícula, frutos retenidos/panícula y redujeron significativamente el pico de caída de frutos 2 semanas después del cuajado. Además, 30 ppm de NAA + 5 ppm de CPPU y 5 ppm de CPPU produjeron el mayor peso de fruto, tamaño de fruto y rendimiento. | [9] |
En conclusión, las citoquininas desempeñan un papel fundamental en el cultivo de aguacate al influir en el crecimiento, la brotación, la floración, la senescencia y la tolerancia al estrés de las plantas. Estas hormonas vegetales pueden estimular el desarrollo vigoroso de la planta, aumentar la producción de flores y frutos, prolongar la vida útil de los frutos después de la cosecha y mejorar la capacidad de las plantas para tolerar el estrés. Sin embargo, es importante tener precaución al aplicar citoquininas, ya que un uso excesivo puede tener efectos negativos. Consultar a expertos en el cultivo de aguacate es recomendable para determinar las prácticas adecuadas de aplicación de citoquininas en un contexto específico. En general, comprender el papel de las citoquininas en el cultivo de aguacate puede ayudar a los agricultores a optimizar el rendimiento y la calidad de los cultivos.
Si quieres incrementar la productividad con nuestros productos, los cuales cuentan con las hormonas vegetales más importantes, puedes compartirnos tus datos y nuestro Ing. Agrónomo más cercano a tu ubicación te contactará muy pronto.
Referencias
[1] C. A. Díaz Diez et al., Actualización tecnológica y buenas prácticas agrícolas (BPA) en el cultivo de aguacate. Corporación Colombiana de Investigación Agropecuaria (Agrosavia), 2020. doi: 10.21930/agrosavia.manual.7403831.
[2] Agronet, “Área, producción y rendimiento nacional por cultivo,” Ministerio de Agricultura y Desarrollo Rural, 2021. https://www.agronet.gov.co/estadistica/Paginas/home.aspx?cod=1 (accessed Apr. 04, 2023).
[3] FAO, “Cultivos y productos de ganaderia,” FAOSTAT, 2023. https://www.fao.org/faostat/es/#data/QCL (accessed Apr. 04, 2023).
[4] A. D’Asaro, C. Reig, A. Martínez-Fuentes, C. Mesejo, V. Farina, and M. Agustí, “Hormonal and carbohydrate control of fruit set in avocado ‘Lamb Hass’. A question of the type of inflorescence?,” Sci Hortic, vol. 282, May 2021, doi: 10.1016/j.scienta.2021.110046.
[5] L. C. Garner and C. J. Lovatt, “Physiological factors affecting flower and fruit abscission of ‘Hass’ avocado,” Sci Hortic, vol. 199, pp. 32–40, Feb. 2016, doi: 10.1016/j.scienta.2015.12.009.
[6] C. S. Moore-Gordon, A. K. Cowan, I. Bertling, C. E. J. Botha, and R. H. M. Cross, “Symplastic Solute Transport and Avocado Fruit Development: A Decline in Cytokinin/ABA Ratio is Related to Appearance of the Hass Small Fruit Variant,” Plant Cell Physiol, vol. 39, no. 10, pp. 1027–1038, Oct. 1998, doi: 10.1093/oxfordjournals.pcp.a029299.
[7] A. K. Cowan, C. S. Moore-Gordon, I. Bertling, and B. N. Wolstenholme, “Metabolic Control of Avocado Fruit Growth (Isoprenoid Growth Regulators and the Reaction Catalyzed by 3-Hydroxy-3-Methylglutaryl Coenzyme A Reductase),” Plant Physiol, vol. 114, no. 2, pp. 511–518, Jun. 1997, doi: 10.1104/pp.114.2.511.
[8] A. K. Cowan and I. D. Railton, “Cytokinins and Ancymidol inhibit Abscisic Acid Biosynthesis in Persea gratissima,” J Plant Physiol, vol. 130, no. 2–3, pp. 273–277, Sep. 1987, doi: 10.1016/S0176-1617(87)80231-6.
[9] L. mostafa, Y. Mostafa, and I. El-Berry, “Effect of NAA and CPPU on Fruit drop, Yield and Quality of Avocado Trees,” Egyptian Journal of Horticulture, vol. 47, no. 2, pp. 137–147, Aug. 2020, doi: 10.21608/ejoh.2020.44228.1144.
